Disclaimer & Copyright Notices; Optimized for the MS Edge
Soil & Water Conservation Society of Metro Halifax (SWCSMH)
Updated: August 03, 2017 ![]()
The productivity of most dystrophic lakes classically has been described as low. However, more detailed examinations indicate the contrary. Phytoplankton biomass (chlorophyll) is significantly higher in coloured ("brown-water") lakes than in clear lakes. Annual productivity of the epilimnetic bacterioplankton was also found to be much higher (four times) in dystrophic lakes than in clear lakes. Anoxic hypolimnia are frequent in dystrophic lakes (cf., Wetzel, 2001).
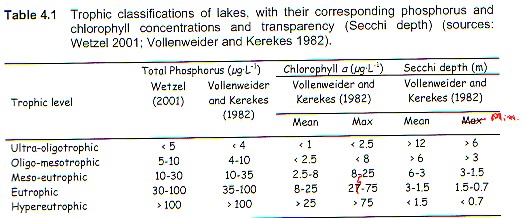
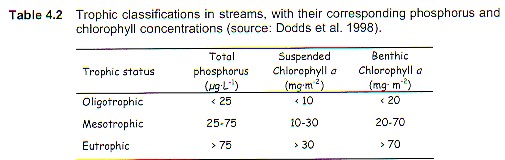
Trophy, then, is an expression of the combined effects of organic matter to the lake! As developed originally and as largely used today, the trophic concept (e.g., TP, Cha, SD, and TN) refers to the pelagic-zone-planktonic portion of the lake ecoststem (cf. Tables 4.1 and 4.2, Environment Canada-2004). The littoral flora and its often dominating supply of autochthonous organic matter to the system, was, and usually still is, ignored.
(Based on organic matter production by phytoplankton and primary physical and chemical determinants; after Naumann, 1929)
Oligotrophy- low production with low P and N
Eutrophy- high production with high P and N
Acidotrophy- low production with low P and N, but pH<5.5
Alkalitrophy- high production with high calcium
Argillotrophy- low production with high clay turbidity
Siderotrophy- low production with high iron
Dystrophy- low production with high humic colour (Thienemann, 1921)

Vollenweider's
dictum- the OECD Probability Distribution Diagrams: Notwithstanding the
trophic state classifications based on pelagic parameters, listen to a 4-minute mp3 sound file of Dr. Richard Vollenweider promoting the use of the OECD probability distribution diagrams in order to establish trophic states with a high confidence level!
Dr. Richard Vollenweider had been recipient of the top medal in limnology, the Naumann-Thienemann Medal, among several other awards!
To avoid erroneous
conclusions concerning trophic state, lakes with high inputs of
allochthonous organic matter (i.e., dystrophic lakes) or lakes where
water color is over 10 pt. units, should not be used for oxygen deficit
calculations. In addition, only lakes with a well-defined thermocline
(>1 �C/m) at the end of the summer stratification are to be
considered, and the hypolimnium is defined as beginning downwards from
the depth of the inflection point during the two months preceding the
onset of the fall overturn. In addition, only lakes where the
hypolimnetic to epilimnetic volume ratio is atleast 1.5 are to be
considered (cf. OECD, Organization for Economic Co-Operation and Development).
There are millions of shallow lakes in the world. Maximum biodiversity of freshwater ecosystems occurs where wetland and littoral habitat heterogeneity interfaces with pelagic regions.
In many shallow lakes and ponds, the littoral structure and its productivity completely dominates the ecosystem.
Loadings of nutrients are proportionally higher, losses are lower, and rates of nutrient cycling are faster in shallow lakes than in deep ones.
In the temperate region, thermal stratification for any appreciable length of time (months) is poor if basin depths are lower than 5 to 7 metres.
Submersed macrophytes dominate, as in large lakes, when nutrient concentration of the water is low (25 �g/l) with a high N:P ratio (>>10:1).
In hardwater, calcareous shallow lakes, charophyte macroalgae are common under oligotrophic conditions.
Isoetid submersed plants (e.g., Is�etes, Littorella, Lobelia) are common in more acidic oligotrophic waters.
Traditionally, limnology is mostly concerned with lakes that stratify in summer. The impact of macrophytes on the community is relatively small in such lakes, as plant growth is restricted to a relatively narrow marginal zone.
These lakes are not expected to stratify for long periods in summer. This type of lake, where the entire water column is frequently mixed, is also referred to as polymictic. The intense sediment-water interaction and the potentially large impact of aquatic vegetation makes the functioning of shallow lakes different from that of their deep counterparts in many aspects.
The pristine state of the majority of shallow lakes is probably one of clear water and a rich aquatic vegetation. Nutrient loading has changed this situation in many cases. The lakes have shifted from clear to turbid, and with the increase in turbidity, submerged plants have largely disappeared.
Restoration of non-vegetated turbid shallow lakes to the clear vegetated state is notoriously difficult. Reduction of the nutrient loading may have little effect, as during the period of eutrophication a large amount of phosphorus has often been adsorbed by the sediment. When the loading is reduced and its concentration in the water drops, phosphorus release from the sediment becomes an important nutrient source for phytoplankton. Thus a reduction of the external loading is often compensated by `internal loading', delaying the response of the lake water concentration to the reduction of external loading.
However, internal loading is not the only reason why restoration of turbid shallow lakes is difficult. With the disappearance of aquatic vegetation the structure of the shallow lake community changes dramatically.
Invertebrates that are associated with vegetation disappear and with these animals the birds and fishes that feed on them or on the plants. Also, vegetation provides an important refuge against predation for many animals, and hence its disappearance causes crucial shifts in many predator-prey relationships. Large zooplankton use vegetation as a daytime refuge against fish predation. In vegetated lakes they can contribute significantly to the control of phytoplankton biomass. In the absence of vegetation their numbers are strongly reduced. This, and the increased nutrient availability, allows phytoplankton biomass to be higher in the absence of vegetation. In addition, wave resuspension of the unprotected sediment can cause a considerable additional turbidity once the vegetation has vanished. The fish community of unvegetated lakes becomes dominated by species that forage on benthic invertebrates. Their activity promotes the nutrient flux from the sediment into the water and causes an extra resuspension of sediment particles, contributing to the already high turbidity.
Ecological feedback mechanisms are thus an important reason why restoration of the vegetated clear water state is difficult. In many cases, nutrient reduction alone may be insufficient to restore the clear state in shallow lakes. Additional measures, however, such as removal of part of the fish stock and changes in the water level, have been successfully used as a way to break the feedback that keeps such lakes turbid.
Until 1980, Little Mere, an artificial lake in North Cheshire, UK, was polluted with effluent from a small sewage treatment works that discharged directly to it. Two consequences of this have been a greater understanding of the link between nutrients and alternative states in shallow lakes, and the socially reprehensible hazards of a large number of impatient people driving cars aggressively on the roads around the lake!
Despite a water chemistry in the lake that resembled a laboratory culture solution for algae, the water of Little Mere was crystal clear in summer and the lake had a substantial coverage of plants. The reason became clear when, one June morning in 1980, it was noticed that the water was full of bright red objects. These turned out to be Daphnia magna, which are full of haemoglobin. This species is so big (up to 4mm) that it cannot coexist with fish, who see it very readily (especially when its haemoglobin content makes it stand out like a bar sign in Las Vegas!).
D. magna was abundant in the lake because the effluent discharge into it was of poor quality and was deoxygenating the water so severely, especially at night, that few or no fish could survive. In turn the Daphnia prospered and effectively prevented any algal growth in the summer.
2.1 Plants can take up large amounts of both nitrogen and phosphorus (luxury consumption) compared with their immediate growth needs.
2.1.1. Except at very low phosphorus concentrations, where charophytes are common and where phosphorus limitation of growth may be important, phosphorus is normally not scarce among dense plant beds, even if luxury uptake occurs, because of release from the sediments. Anaerobic sediments favour phosphate release and the sediments laid down under plant beds tend to be deficient in oxygen through decomposition of sloughed off leaves and periphyton and the excreta of the invertebrate populations. Even aerobic sediments in shallow lakes emit much phosphorus, however, perhaps by physico-chemical as well as biologically mediated mechanisms.
2.1.2. Combined nitrogen, however is very vulnerable. Not only can it be taken up by the plants, it can be denitrified to nitrogen gas especially in the conditions within plant beds, which include a gradient from severe deoxygenation at night to supersaturation with oxygen at the surface by day.
2.1.2.1. Such alternation favours mineralisation of combined nitrogen to ammonia and oxidation to nitrate followed by denitrification to nitrogen gas. Combined nitrogen is frequently undetectable in the open water of plant-dominated lakes during the summer.
3.1. Plant photosynthesis changes the chemistry of the water. Carbon dioxide and bicarbonate are withdrawn and through the operation of well-known carbon equilibria, this increases the pH often to values well above 9, sometimes 10. This appears to inhibit fish feeding and provides a chemical refuge in addition to the deoxygenation deeper in the beds caused by decomposition of organic matter in the darkness of the lower layers.
4.1. The ratio of piscivorous fish biomass to zooplanktivorous fish biomass tends to be higher in plant-dominated lakes so that annual recruitment to the populations of the latter may be restricted by predation. These populations include the young of plant-invertebrate-eating fish.
4.2. Fish need sufficient light and sufficient striking distance to capture their prey and, for the most part, dense plant beds frustrate these requirements. Some fish such as perch (Perca fluviatilis) cope better than others like roach (Rutilus rutilus) but experiments have shown that when the density of plants is reduced, perch are more readily able to feed on Daphnia. Daphnia buffers are most important in the early establishment of plants but may become less important than nitrogen limitation once the plant community has fully developed.
5.1. Periphyton algae may pose as great a competitive threat to submerged plants as phytoplankton algae. They may form a dense fur over the surfaces and compete for carbon dioxide, light and nutrients.
5.1.1. Periphyton growths are often controlled by the grazing of macroinvertebrates such as snails, mayfly nymphs and chironomid larvae.
6.1. Evidence of allelopathy is strong in the laboratory, but lacking in the field. Plants do secrete substances which inhibit growth of laboratory algal cultures and some of the substances have been characterised. However, the extent to which these substances, faced with a barrage of heterotrophic bacteria, may accumulate and be effective in natural waters on a range of many species of algae which may easily substitute for one another, is unknown and will be extremely difficult to investigate. The potential, however, undoubtedly exists.
5.1. The sediment laid down by phytoplankters is more fluid and amorphous than that laid down in plant beds. It may not provide a firm rooting medium for plants, and it may be more readily disturbed to give inorganic turbidity in the water and an unfavourable light climate for plant development.
6.1. The impact of fish predation on a zooplankton community that has developed in the previous absence or scarcity of fish is to convert a community dominated often by large Cladocera, or sometimes a mixture of Cladocera and copepods to one dominated by rotifers and copepods, with perhaps some of the smaller cladoceran genera. The former community is efficient at grazing and can keep the water clear, the latter community is relatively inefficient and associated algal populations can quickly build up.
![]()

We salute the Chebucto Community Net (CCN) of Halifax, Nova Scotia, Canada for hosting our web site, and we applaud its volunteers for their devotion in making `CCN' the best community net in the world

{kind=link}
{kind=link}
{kind=link}
{kind=link}